
Trichoptera
EOL Text
Order Trichoptera have membranous wings and long antennae. They are attracted to blue lights, which are frequently used to collect them. Order Trichoptera undergo complete metamorphosis. The larvae resemble caterpillars and are aquatic. The larvae either create nets or cases underwater. They feed off of prey trapped in the nets or cases. Cases or nets use twigs, pebbles, leaves, sand, and other small bio matter. The materials are glued together with a special silk. The cases or nets are then cemented to an underwater object, like a river rock. If the larvae do not use cases or nets, they are predaceous. The larvae with cases pupate inside their case for a few weeks to a few months and emerge fully developed. The larval stage usually lasts for about a year and the adults will live for about a month. Caddisflies are found in the fossil record as far back as the Triassic.
| License | http://creativecommons.org/licenses/by/3.0/ |
| Rights holder/Author | Rhianna Hruska, Rhianna Hruska |
| Source | No source database. |
Trichoptera, or caddisflies, comprise the most diverse insect order whose members are exclusively aquatic. Only aquatic Diptera outnumber them in species and ecological diversity. The larval stages are found in lakes, rivers, and streams around the world, and are important components of food webs in these freshwater ecosystems (Resh and Rosenberg 1984). A few species in the family Chathamiidae from New Zealand and Australia are unusual for the Insecta in having larvae that are truly marine, mostly restricted to tidal pools.
Adult Trichoptera, in contrast to the larvae, are terrestrial and look much like drab, fragile moths, often occurring in large numbers in lakeside or streamside habitats. The similarity is not incidental. Trichoptera are closely related to the order Lepidoptera and together the two orders comprise the superorder Amphiesmenoptera, or "dressed-up wings," the name referring to the dense clothing of scales or hairs on the wings. Monophyly of these two orders is strongly supported in both morphological and molecular analyses (Kristensen 1984, Wheeler, et al. 2001). Trichoptera possess the more primitive character state, having hairs rather than scales, and this character accounts for the name Trichoptera, meaning "hairy wings." Also, unlike moths and butterflies, which typically have a coiled, tubelike proboscis for feeding, adult caddisflies lack well-developed mouthparts, including the absence of mandibles, but have a well-developed haustellum (synapomorphic for the order) formed from a fusion of the hypopharynx and labium, and used in some species to imbibe liquids.
Unlike Lepidoptera larvae, which are predominantly terrestrial herbivores, Trichoptera larvae, with very few exceptions, are aquatic and primarily detritivorous. Like lepidopteran caterpillars, caddisfly larvae are capable of spinning silk from specially modified salivary glands. The diversity of microhabitats exploited by caddisfly larvae is a consequence of the many ways silk is used to construct retreats, nets, and cases and probably accounts for the success of the order as a whole (Mackay and Wiggins 1979, Wiggins 1996).
Almost 12,000 caddisfly species, placed in 45 families and about 600 genera, have been described from all faunal regions, but it has been estimated that the world fauna may contain as many as 50,000 species (Schmid 1984). The three currently recognized suborders are largely characterized by differences in the way silk is used (Ross 1944), whether to produce nets or tubes, or as glue to make various types of portable cases, often incorporating sand and small pebbles, or bits of leaves and twigs, each genus or even species building its own particular style of case. Some larvae are free-living and predaceous, but nevertheless lay down a strand of silk as they move, much like the larvae of Lepidoptera.
The larvae, and the fascinating nets and cases they produce, represents the life stage most familiar to the non-entomologist, and the case-making behavior of some species may account for the English common name, caddisfly. Although the origin of the word is obscure, it has been suggested to derive from cadaz or cadace (caddys), a word of variable spelling used in Shakespearean times to refer to a ribbon made from a certain kind of yarn sold by traveling vendors, who because of this were sometimes called "cadice men." Cadice men would pin samples of their wares to their clothing, a habit which may have suggested the name caddisfly or caddisworm for the aquatic larvae, who exhibit the analogous behavior of attaching bits of leaves and twigs to the outside of their cases (Hickin 1967).
Although caddisflies are not generally considered to be of great economic importance as pests, they are beneficially important in the trophic dynamics and energy flow in aquatic ecosystems. The larvae are also useful as biological indicator organisms for assessing water quality. Extensive use of them has been made for this purpose because larvae of different species vary in sensitivity to various types of pollution (Resh and Unzicker 1975, Resh 1993, Dohet 2002), and because the taxonomy of the group is relatively well known for temperate regions. Unfortunately, the larvae of many species, especially in the tropics, are unknown or have not been associated with their adult forms.
| License | http://creativecommons.org/licenses/by-nc-sa/3.0/ |
| Rights holder/Author | Ralph W. Holzenthal, Roger J. Blahnik, Aysha Prather, Karl Kjer, Tree of Life web project |
| Source | http://tolweb.org/Trichoptera/8230 |
Caddisflies are easily recognized by a number of features. Adult mouthparts are reduced, with mandibles essentially absent, but the maxillary and labial palps, and often the haustellum, are prominent. The compound eyes are well developed, and ocelli may or may not be present. The forewings are somewhat longer than the hind wings, although the hind wings may be broader. Both pairs of wings and the body are usually covered with setae, or hairs, and occasionally with patches of scales. The wings are generally held rooflike when folded over the body. In most species the antennae are filamentous and about as long as the body, but in some families they can be several times longer than the body. Tibial spurs on the legs are conspicuous.
Larvae are aquatic and construct a case or retreat, except for a couple of "free-living" families. Like most holometabolous larvae, they have well developed mandibulate mouthparts and the thoracic legs are well developed, but abdominal prolegs are absent except for a pair of anal prolegs on the last abdominal segment, each proleg bearing a strong "anal claw." The exarate pupae are also aquatic, and have dectitious mandibles in most families.

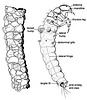
Left: Caddisfly adult (Spicipalpia: Hydrobiosidae). Illustration copyright © 2004, Lourdes Chamorro-Lacayo. Right: Caddisfly larva and case (Integripalpia: Odontoceridae). Illustration copyright © 2004, Lourdes Chamorro-Lacayo.
The monophyly of the order Trichoptera is very well established. Following is a list of some characters that have been proposed as synapomorphic for the order (Weaver 1984, Kristensen 1991):
- Larvae aquatic, apneustic (no open spiracles), respiration epidermal, often by filamentous abdominal gills
- Larval tentorium reduced, delicate
- Larval antennae greatly reduced
- Larval abdominal segments I-IX without prolegs
- Larval abdominal segment IX with dorsal tergite
- Adult mandibles reduced, with loss of mandibular articulation
- Adult prelabium joined with hypopharynx to form a unique "haustellum" which serves as a lapping/sponging organ
| License | http://creativecommons.org/licenses/by-nc-sa/3.0/ |
| Rights holder/Author | Ralph W. Holzenthal, Roger J. Blahnik, Aysha Prather, Karl Kjer, Tree of Life web project |
| Source | http://tolweb.org/Trichoptera/8230 |
Animal / parasite / endoparasite
cyst of Dolichosaccus rastellus endoparasitises larva of Trichoptera
Animal / pathogen
pure white to grey or rarely green, shaggy rhizoids of Erynia conica infects adult of Trichoptera
In Great Britain and/or Ireland:
Animal / pathogen
Erynia rhizospora infects adult of Trichoptera
Animal / parasite / endoparasite
cyst of Opisthioglyphe ranae endoparasitises larva of Trichoptera
| License | http://creativecommons.org/licenses/by-nc-sa/3.0/ |
| Rights holder/Author | BioImages, BioImages - the Virtual Fieldguide (UK) |
| Source | http://www.bioimages.org.uk/html/Trichoptera.htm |
Trichoptera (Trichoptera 10 families 21 spp.) is prey of:
Serranochromis robustus
Barbus paludinosus
Gasterosteus aculeatus
Salvelinus fontinalis
Eleutherodactylus coqui
Eleutherodactylus portoricensis
Anolis evermanni
Anolis gundlachi
Sphaerodactylus klauberi
Odonata
Gonatista grisea
Based on studies in:
Malawi (River)
Africa, Crocodile Creek, Lake Nyasa (Lake or pond)
England, River Cam (River)
Canada: Ontario, Mad River (River)
Puerto Rico, El Verde (Rainforest)
This list may not be complete but is based on published studies.
- G. Fryer, The trophic interrelationships and ecology of some littoral communities of Lake Nyasa, Proc. London Zool. Soc. 132:153-229, from p. 219 (1959).
- G. Fryer, 1957. The trophic interrelationships and ecology of some littoral communities of Lake Nyasa with special reference to the fishes, and a discussion of the evolution of a group of rock-frequenting Cichlidae. Proc. Zool. Soc. London 132:153-281, f
- W. E. Ricker, 1934. An ecological classification of certain Ontario streams. Univ. Toronto Studies, Biol. Serv. 37, Publ. Ontario Fish. Res. Lab. 49:7-114, from pp. 78, 89.
- P. H. T. Hartley, Food and feeding relationships in a community of fresh-water fishes, J. Anim. Ecol. 17(1):1-14, from p. 12 (1948).
- Waide RB, Reagan WB (eds) (1996) The food web of a tropical rainforest. University of Chicago Press, Chicago
| License | http://creativecommons.org/licenses/by/3.0/ |
| Rights holder/Author | Cynthia Sims Parr, Joel Sachs, SPIRE |
| Source | http://spire.umbc.edu/fwc/ |
Trichoptera (Trichoptera 10 families 21 spp.) preys on:
algae
detritus
plant fragments
Bacillariophyceae
Coscinodiscus
Synedra
Based on studies in:
Malawi (River)
Africa, Crocodile Creek, Lake Nyasa (Lake or pond)
England, River Cam (River)
This list may not be complete but is based on published studies.
- G. Fryer, The trophic interrelationships and ecology of some littoral communities of Lake Nyasa, Proc. London Zool. Soc. 132:153-229, from p. 219 (1959).
- G. Fryer, 1957. The trophic interrelationships and ecology of some littoral communities of Lake Nyasa with special reference to the fishes, and a discussion of the evolution of a group of rock-frequenting Cichlidae. Proc. Zool. Soc. London 132:153-281, f
- P. H. T. Hartley, Food and feeding relationships in a community of fresh-water fishes, J. Anim. Ecol. 17(1):1-14, from p. 12 (1948).
| License | http://creativecommons.org/licenses/by/3.0/ |
| Rights holder/Author | Cynthia Sims Parr, Joel Sachs, SPIRE |
| Source | http://spire.umbc.edu/fwc/ |
There has been considerable disagreement about the basal relationships of Trichoptera. This has resulted not only in different hypotheses about the evolutionary history of the group, but also in a confusion in the use of taxonomic categories, since different authors use different terminology, or have been inconsistent in how certain taxonomic categories have been used. In general, three major groups have been recognized, more or less corresponding to the different ecological adaptations of the larvae. We refer to these by their most common subordinal names, Annulipalpia, Spicipalpia, and Integripalpia (each in its most restricted sense and as used by Wiggins and Wichard 1989). However, the respective superordinal names of Hydropsychoidea, Rhyacophiloidea, and Limnephiloidea, respectively (sensu Ross 1956, Neboiss 1991), have sometimes been used to refer to groups of equivalent taxonomic coverage.
Because life history strategies have been used in constructing theories of caddisfly phylogenetic relationships, it is useful to review the life histories of the major groups:
- The Annulipalpia includes all of the families whose larvae make retreats and capture nets.
- The Spicipalpia includes several rather different groups, each with different larval habits:
- free-living and predaceous (Rhyacophilidae and Hydrobiosidae) - build no larval structures, but pupate within a domelike enclosure of mineral fragments.
- purse-case makers (Hydroptilidae) - free-living until the last larval instar, and then construct a case which is portable or cemented to the substrate and in which the larvae eventually pupate.
- tortoise-case or saddle-case makers (Glossosomatidae) - make a case that is constructed very much like the pupal cases of Annulipalpia and the free-living Spicipalpia, consisting of a dome of small sand grains and pebbles. However (to complete the analogy to a tortoise) the larva also makes a transverse strap beneath the dome, allowing the larva to carry it on its back. The tortoise-case makers construct a new and bigger case with each larval instar, and then pupate within the last larval case, after removing the ventral strap and attaching the case to the substrate.
- The Integripalpia have been called the tube-case makers, because they most commonly construct a tubular case. The case, however, can be made from very different materials or formed in peculiar ways in various species. The larvae are mobile and simply extend or add to the case with each larval instar, eventually pupating inside the slightly modified larval case.
There is broad agreement about the monophyly of two of these major taxonomic groups, the Annulipalpia and Integripalpia (as defined above), and considerable disagreement about the monophyly and relative placement of taxa within the Spicipalpia.
Morse (1997) provided a thorough summary of the hypotheses of relationships proposed for the major groups of Trichoptera and also of the status of phylogenetic work at the family and genus level undertaken to date. Ross (1956) was the first to identify explicitly any derived characters for caddisfly taxa. He recognized two monophyletic suborders, Annulipalpia (equivalent to his superfamily Hydropsychoidea) and Integripalpia. These subordinal names were first established by Martynov (1924). Ross's Integripalpia contained two superfamilies, Rhyacophiloidea and Limnephiloidea. These superfamilies are equivalent to the suborders Spicipalpia and Integripalpia, as used here. However, Ross's Rhyacophiloidea (Spicipalpia) was paraphyletic as originally defined. Further, his hypothesis of the relationships among the three rhyacophiloid families (Rhyacophilidae [including Hydrobiosidae], Glossosomatidae, and Hydroptilidae) and the Limnephiloidea was based mainly on a presumed evolutionary transformation in larval case/pupal cocoon making behavior. Ross lacked morphological synapomorphies at his Glossosomatidae + Hydroptilidae + Limnephiloidea node and had only one at his Hydroptilidae + Limnephiloidea node. Thus his behavioral transformation series is supported by only one non-behavioral character.

Ross (1956, 1964, 1967)
Weaver (1983, 1984, 1992a,b; also Weaver & Morse, 1986) was the first to apply cladistic principles to caddisfly higher level classification and examined about twice as many morphological characters as Ross. Weaver (1984) concluded that Spicipalpia (which he treated as an infraorder within a more broadly defined suborder Annulipalpia) was monophyletic and had a sister taxon relationship to the infraorder Curvipalpia (= Annulipalpia of Ross and as used here). Weaver restricted his concept of Integripalpia to include only the Limnephiloidea of Ross, and this is the sense in which it is used here.


Left: Tree of Weaver (1984). Right: Tree of Wiggins and Wichard (1989)
Later, Wiggins and Wichard (1989; also Wiggins 1992, Wichard 1991, Wichard et al. 1993, Wichard et al. 1997) proposed a sister group relationship between Annulipalpia and Integripalpia, based on an interpretation of pupal cocoon evolution in Trichoptera. Their phylogeny is based on the hypothesis that the closed, semipermeable cocoons of parchmentlike silk found in the spicipalpian families represent the groundplan condition of the order, and the cocoons of permeable silk with ventilation openings found in the Annulipalpia and Integripalpia are derived. A more detailed account of behavioral evolution in Trichoptera is found in the treatise of Frania and Wiggins (1997), who hypothesized that the ancestral habitat for the order Trichoptera was in cool, flowing, well-oxygenated water. This can be contrasted with the theory of Weaver and Morse (1986), who proposed that the ancestral trichopteran habitat was in subterranean silk-lined tubes in saturated soil. However, Kristensen (1997), pointed out that silk-lined tubes in Lepidoptera do not appear until the eight branch from the base of the tree, in Neolepidoptera (Exoporia), and thus this adaptation cannot be assumed to be ancestral within Trichoptera. Based on their hypothesis of evolution in the group, Wiggins, Wichard, and Frania would elevate Weaver's Infraorder Spicipalpia to a third suborder, coordinate with Annulipalpia and Integripalpia. According to Wiggins (1992), the pupation hypothesis was not intended as a statement of higher level trichopteran relationships; the recognition of Spicipalpia was one of convenience, since it serves to focus discussion on the unresolved problem of basal relationships in Trichoptera. These workers preferred to consider the relationships of the three suborders as currently unresolved.
In the study by Frania and Wiggins (1997), the hypothesis of a basal Spicipalpia was tested using a computer assisted parsimony analysis of 70 morphological larval and adult characters, which included the majority of families in the order. Characters were polarized using a hypothetical caddisfly ancestor, whose character states were inferred through consideration of character states in Mecoptera and Lepidoptera. The results could be considered equivocal at best, since they do not support either monophyly of Spicipalpia or of a basal position of Spicipalpia. The strict consensus phylogeny most closely resembles the hypothesis of Ivanov (1997, 2002):
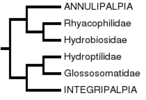
Ivanov (1997, 2002)
Ivanov proposed that Hydroptilidae and Glossosomatidae of the Spicipalpia are sister taxa and allied to the Integripalpia, and that Rhyacophilidae and Hydrobiosidae are sister taxa allied to the Annulipalpia. He specifically challenged Weaver's hypothesis of spicipalpian monophyly, providing evidence that each of Weaver's 4 spicipalpian apomorphies are plesiomorphic. Frania and Wiggins similarly found Hydroptilidae to be closely related to Integripalpia, but the relationship of Glossosomatidae was less certain, depending on the analysis performed. Using selected characters they suggested that Glossosomatidae might be allied to Hydrobiosidae and Rhyacophilidae and basal to Annulipalpia, but were uncertain about the relationship of Hydroptilidae. In the cladograms presented, it was sister taxon to the Integripalpia, suggesting that Spicipalpia may not be monophyletic.
The most recent and complete analysis of basal relationships in Trichoptera is that of Kjer et al. (2001, 2002), who used a molecular dataset of several gene fragments, including mitochondrial, and nuclear DNA, and also included the morphological characters of Frania and Wiggins. Forty-three of forty-five families were included, and both parsimony and likelihood analyses were performed. This combined data analysis is the hypothesis of relationships presented above. In Kjer et al.'s analysis, Annulipalpia and Integripalpia were monophyletic and Spicipalpia was paraphyletically arranged at the base of the Integripalpia, as in Ross's hypothesis. The relationship of spicipalpians to Integripalpia was strongly supported, and Kjer et al. (2001, 2002) rejected the separation of some families of spicipalpians with Annulipalpia, as in the Ivanov (1997, 2002) and Frania and Wiggins (1997) morphology-based hypotheses. In fact, a differentially weighted analysis performed in Kjer et al. (2001) of the Frania and Wiggins (1997) morphological data recovered a phylogenetic hypothesis that was identical to Ross (1967). However, paraphyly of Spicipalpia was only weakly supported and the possibility that Spicipalpia is monophyletic could not be eliminated. Some partitions of the data supported a monophyletic Spicipalpia, and by most analytical criteria, a monophyletic Spicipalpia was only slightly suboptimal.
Determining the relative relationships among the families of Spicipalpia remains the next challenging question in resolving basal relationships in Trichoptera and in helping to resolve the question of the evolution of net and case-making behavior in the order.
| License | http://creativecommons.org/licenses/by-nc-sa/3.0/ |
| Rights holder/Author | Ralph W. Holzenthal, Roger J. Blahnik, Aysha Prather, Karl Kjer, Tree of Life web project |
| Source | http://tolweb.org/Trichoptera/8230 |
Portable cases protect from predators: caddisflies
Some caddisflies protect themselves from predators by building portable cases out of local materials - such as pebbles, sand, and aquatic plants - that are cemented together with silk or mucus.
"There are a number of invertebrates (animals without backbones) that construct cases for themselves from materials abundant in their environments, like sand, pebbles, shells, and plant materials. These bits and pieces are cemented together with silky or mucous secretions from the creature. Caddis flies and sabellid or honeycomb tubeworms are two common examples. Whereas caddis flies appear to randomly select objects of varying sizes, honeycomb tubeworms select particles of uniformly small size for the primary layer of their tubes." (Biomimicry Guild unpublished report)
Learn more about this functional adaptation.
| License | http://creativecommons.org/licenses/by-nc/3.0/ |
| Rights holder/Author | (c) 2008-2009 The Biomimicry Institute |
| Source | http://www.asknature.org/strategy/ed570c884d6db0d32744ba62a10e0578 |
Barcode of Life Data Systems (BOLD) Stats
Specimen Records:58626
Specimens with Sequences:51496
Specimens with Barcodes:42806
Species:5835
Species With Barcodes:5153
Public Records:39405
Public Species:1928
Public BINs:5190